


Insana Lab: Ultrasonic Imaging - The University of Illinois at Urbana-Champaign
Project 4: Atherosclerosis and the role of cell adhesion and blood flow dynamics
Jean K. Tsou [jktsou.ucdavis.edu]
|
|
|
|
|
|
|
|
|
|
|
|
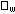 decreases linearly across the flow chamber.
Along the center of the flow chamber the shear stress is a function of viscosity decreases linearly across the flow chamber.
Along the center of the flow chamber the shear stress is a function of viscosity 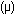 ,
volumetric flow rate (Q), channel height (h = 100 ,
volumetric flow rate (Q), channel height (h = 100 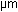 ), entrance width (w1 = 2 mm),
channel length (L) and position (x). ), entrance width (w1 = 2 mm),
channel length (L) and position (x).
Fig. 4b shows the setup of the in vitro flow experiments on human aortic endothelial cells (HAECs). The cell cover slip is temporarily bound to the PDMS flow chamber by vacuum. Flow medium was infused into the chamber by a syringe pump to create a known, spatially varying shear field. |
|
|
|
|
|
|
|
|
References:
[1] AM Malek, S. A., S Izumo (1999). "Hemodyamic shear stress and its role in atherosclerosis." JAMA 282: 2035-2042. |